Publié le 23 juin 2019 et mis à jour le 17 octobre 2024.
La vie en société chez les insectes
Eusocialité, évolution et sélection de parentèle
Le nombre d’espèces sociales est faible, mais leur biomasse, la masse totale de tous les individus de ces espèces, est bien loin d’être négligeable. La socialité est sans aucun doutes la clé du succès de nombreux insectes. Les termites, guêpes, frelons, bourdons, certaines abeilles comme Apis mellifera et Apis cerana, et bien sûr les fourmis, sont établis sur la plupart des continents et dans la plupart des écosystèmes. Souvent moins connus, d’autres arthropodes vivent aussi en sociétés (des araignées et des crevettes) et il existe même un vertébré formant des colonies, le rat-taupe nu. Chez tous ces animaux, la plupart des membres de la colonie ne se reproduisent jamais dans leur vie, et se consacrent entièrement à aider les quelques individus qui peuvent se reproduire. Ces animaux sont dits eusociaux. Entre les animaux qui vivent complètement seuls et ceux qui vivent en sociétés, il existe un peu tous les niveaux.

Quels sont les différents niveaux de vie en société ?
Beaucoup d’espèces animales sont solitaires et n’interagissent avec leurs semblables que très peu, accidentellement ou juste en période de reproduction. C’est aussi vrai chez les insectes et la plupart des arthropodes.
Certains animaux se regroupent cependant en agrégats, c’est à dire réunis autour d’une même source de nourriture par exemple, mais ils n’interagissent pas ou peu. Par exemple, certains papillons se posent ensemble sur les flaques d’eau, ou certains coléoptères se nourrissent sur un même cadavre, sans interagir. Se rassembler en agrégats peut cependant avoir des avantages, par exemple les chances d’être choisi comme proie par un prédateur peuvent être plus faibles lorsque vous êtes entourés de centaines de semblables, mais peut aussi augmenter la compétition pour les ressources [1].

D’autres espèces d’insectes sont grégaires, cela signifie qu’elles intéragissent durant certaines activités et préfèrent vivre en groupes [2]. C’est par exemple le cas de certaines blattes, qui émettent des phéromones de regroupement [3] et choisissent leurs abris en fonction du nombre de blattes qui y sont déjà installées [4].
Certains insectes sont qualifiés de subsociaux, c’est le cas par exemple des forficules ou perce-oreilles et des bousiers, qui ont des comportements de soins parentaux (ils prennent soin des oeufs, parfois même des jeunes) [5]. D’autres arthropodes, les mille-pattes polydesmidés par exemple, construisent des "nids" pour leurs oeufs et les scolopendres femelles les gardent avec elles jusqu’à l’éclosion, entre leurs pattes [6].

D’autres sont coloniaux cela signifie qu’ils partagent un même site d’élevage des jeunes, ou communaux, lorsqu’ils partagent un même site d’élevage et que les adultes coopèrent [7]. Les animaux eusociaux "primitifs" élevent leurs jeunes communément et certains sont spécialisés dans certaines tâches. Par exemple certains adultes cherchent à manger et d’autres prennent soin des jeunes. Ces comportements sont communs chez les oiseaux et mammifères marins mais peu représentés chez les insectes. On les observe parfois chez certaines espèces d’abeilles [8]. Un cas particulier et difficile à classer est celui de certaines araignées qui coopèrent durant la chasse ou la construction des toiles [9].
Enfin, les animaux eusociaux dits "évolués" [10] sont ceux chez qui la spécialisation des tâches est très marquée, puisque certains individus se reproduisent tandis que d’autres individus ne se reproduisent pas directement et s’occupent principalement des reproducteurs ou de la colonie. Pour cette raison, les animaux eusociaux sont parfois présentés comme des super-organismes où chaque individu correspondrait à une cellule spécialisée dans une fonction particulière (reproduction, fourragement, circulation dans la colonie, soutien des jeunes individus) [11].

Tableau 1 : Les différents niveaux de socialité.
| Solitaires | Solitaire pour toute activité |
| Agrégats | Réunis autour d’un même stimulus, sans interactions |
| Grégaires | Interactions entre individus (pour au moins une jusqu’Ã la plupart des activités) |
| Subsociaux | Comportements parentaux |
| Coloniaux | Sites d’élevage commun |
| Communaux | Coopération dans les soins aux jeunes |
| Eusociaux "primitifs" | Elevage commun, spécialisation des tâches |
| Eusociaux "évolués" | Spécialisation de la reproduction |
Donc certaines espèces sont sociales et d’autres non ?
Malheureusement ce n’est pas aussi simple. Le degré de socialité peut évoluer durant la vie : les jeunes épeires diadèmes coopèrent à la naissance, mais les adultes se dévoreraient s’ils se rencontraient en dehors des périodes de reproduction [12] [13] !
De la même façon, certains criquets vivent en groupes durant une période de leur vie, et sont solitaires le reste du temps [14]. Il apparaît même parfois dans les colonies d’abeilles des ouvrières qualifiées d’anarchistes par les scientifiques, qui se mettent à pondre des oeufs et à utiliser les resources de la colonies pour leur propre intérêt malgré la présence de la reine [15].
Un cas particulier compliqué est celui de certaines guêpes, chez qui certaines larves se développent comme des soldates qui ne deviendront jamais adultes mais protègent les autres larves. Les larves pourraient donc être qualifiées d’eusociales mais les adultes sont solitaires [16].
Comment la vie en société a-t-elle évoluée chez les animaux ? Pourquoi certains animaux vivent en colonies ?
La vie en société est apparue plusieurs fois au cours de l’évolution. Les fourmis, les abeilles sociales, les bourdons, les guêpes, les frelons et les termites sont des insectes eusociaux parfois très éloignés sur le plan de l’évolution. Ce ne sont pas les seuls arthropodes eusociaux, moins connues, il existe des crevettes eusociales qui vivent en colonies organisées de manière similaire aux colonies de fourmis [17].
Chez les vertébrés, les rat-taupes sont un exemple d’eusocialité : ces rongeurs vivent sous-terre et seul une femelle par colonie se reproduit.
Chez les animaux vertébrés qui vivent en groupe, les causes de la formation des groupes sont souvent la nécessité d’une défense commune contre les prédateurs (effet de dilution, de confusion ou partage de la vigilance) ou au contraire le besoin de coopérer durant la recherche de nourriture (prédation de plus grosses proies, détection de la nourriture facilitée) [18]. Chez les insectes eusociaux, la construction du nid, la défense collective et le fourragement sont aussi des parties importantes de la vie en groupe qui ont pu favoriser l’évolution de l’eusocialité [19].

Historiquement, la théorie de sélection de parentèle [20] [21] propose d’expliquer pourquoi certains individus "acceptent" de ne pas se reproduire tout en permettant à d’autres mebres de la colonie de le faire.
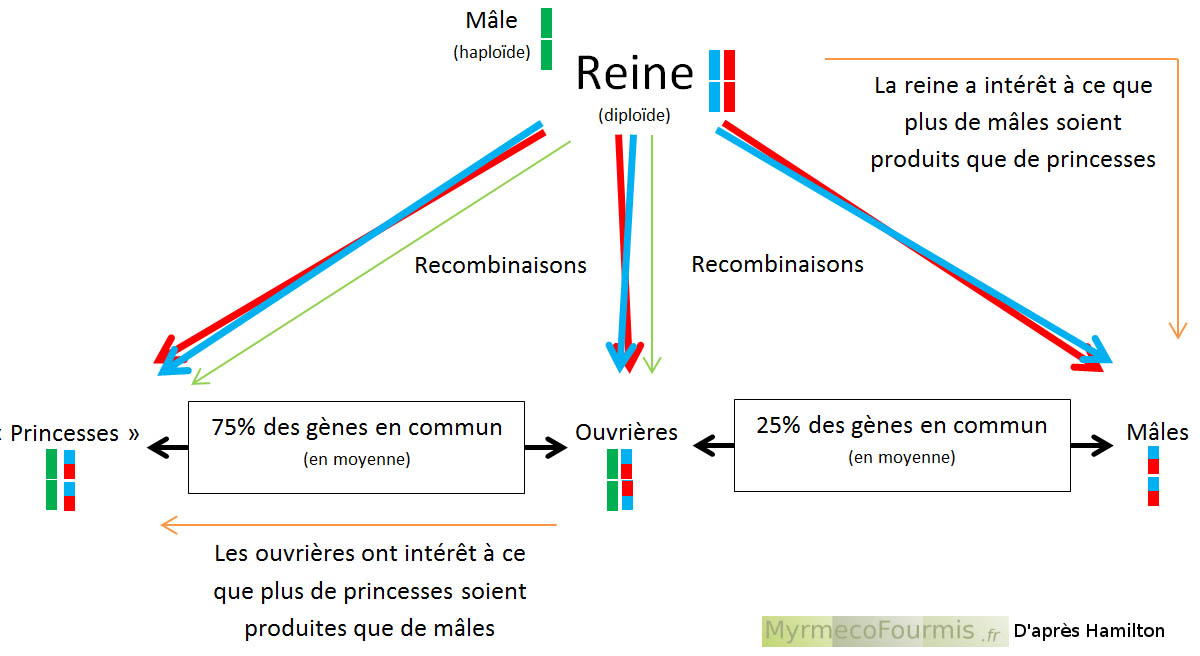
La théorie de sélection de parentèle d’Hamilton se base sur le fait que les individus non reproducteurs (ouvrières) ont un lien génétique très fort avec les individus reproducteurs (les reines). Il peut être plus efficace, en terme de transmission du patrimoine génétique, d’aider une de ses soeurs "reine" à se reproduire plutôt que de se reproduire soi-même. Une ouvrière a dans ce cas intérêt à vivre en société et à se consacrer entièrement à la production de ses soeurs "reines". Hamilton a ainsi établit une formule permettant de comprendre si un animal a ou non intérêt à vivre en société. Pour qu’une vie eusociale soit bénéfique pour une ouvrière, il faut que les liens de parenté (r) et le bénéfice tiré de la vie en société par les individus reproducteurs (b) soient supérieurs aux coûts de la vie en société pour l’ouvrière (c), soit r * b - c > 0 .
Cette théorie est aujourd’hui très critiquée pour des raisons assez subtiles sur lesquelles il serait compliqué de s’étendre ici. D’autres modèles souvent plus complexes la complémentent ou remplacent aujourd’hui [19] [22].
Comment le travail est-il divisé au sein des colonies ?
La division sociale du travail peut prendre deux aspects chez les insectes eusociaux. Elle peut se faire en fonction de l’âge, on parle alors de polyéthisme d’âges. Elle peut aussi se faire en fonction de la morphologie, on parle alors de polyéthisme de castes (mâles, femelles reproductrices et non reproductrices) et de sous-castes (soldates, ouvrières, fourmis pot-à-miel,...), ou de polyphénisme. Bien souvent, le polyphénisme est associé à un polyéthisme d’âges.
Chez les fourmis coupeuses de feuilles du genre Acromyrmex, les grandes ouvrières sont souvent spécialisées dans le transport de fragments de feuilles, alors que les petites sont parfois spécialisées dans la défense des grandes contre de minuscules mouches parasites de fourmis !

Chez certaines abeilles sociales et fourmis, le polyéthisme d’âge commence par une spécialisation dans les tâches internes au nid [23]. Les jeunes fourmis s’occupent donc du couvain, lèchent les oeufs et nourrissent les larves. Puis, elles participent aux travaux d’aménagement et d’agrandissement du nid (excavation pour les fourmis, construction de cire pour les abeilles). Elles montent ensuite la garde à l’entrée, puis en vieillissant elles partent explorer et chercher de la nourriture hors du nid. Enfin, les ouvrières les plus vieilles s’occupent d’éloigner les déchets et les cadavres de la colonie. De cette manière, ce sont toujours les ouvrières les plus âgées qui sont exposées aux risques de prédation ou de contamination, ce qui permettrait de minimiser les pertes pour la colonie [24].
Cet article est agrégé au site du Café des Sciences.
Articles liés
-
 Comment reconnaître les cellules royales, d’ouvrières, de faux-bourdons et les alvéoles de miel et pollen dans une ruche
Comment reconnaître les cellules royales, d’ouvrières, de faux-bourdons et les alvéoles de miel et pollen dans une ruche Reconnaître les alvéoles d’une ruche permet d’anticiper l’état de la colonie. Voici un guide rapide pour permettre aux apiculteurs et apicultrices d’identifier facilement les cellules royales, les cellules de faux-bourdons ou d’ouvrières ainsi que celles contenant du pollen ou du miel.
-
 Que mange le gendarme (l’insecte) ?
Que mange le gendarme (l’insecte) ? Les punaises gendarmes (Pyrrhocoris apterus) sont des insectes rouges et noirs fréquents dans les jardins potagers. Que mangent-ils ? Sont-ils nuisibles ? Sont-ils dangereux ? Toutes les réponses sont ici, avec un petit aperçu de leur biologie.
-
 Chasser les fourmis de la maison
Chasser les fourmis de la maison Comment se débarrasser des fourmis qui rentrent dans la maison ? voici des conseils et astuces simples qui marchent vraiment pour éviter les problèmes de fourmis dans les maisons et appartements.
Notes et références
[1] Parrish, J. K., & Edelstein-Keshet, L. (1999). Complexity, pattern, and evolutionary trade-offs in animal aggregation. Science, 284(5411), 99-101.
[2] Cannings, C., & Orive, L. M. C. (1975). On the adjustment of the sex ratio and the gregarious behaviour of animal populations. Journal of theoretical biology, 55(1), 115-136.
[3] Bell, W. J., Parsons, C., & Martinko, E. A. (1972). Cockroach aggregation pheromones : analysis of aggregation tendency and species specificity (Orthoptera : Blattidae). Journal of the Kansas Entomological Society, 414-421.
[4] Jeanson, R., Rivault, C., Deneubourg, J. L., Blanco, S., Fournier, R., Jost, C., & Theraulaz, G. (2005). Self-organized aggregation in cockroaches. Animal behaviour, 69(1), 169-180.
[5] Tallamy, D. W. (1984). Insect parental care. Bioscience, 34(1), 20-24.
[6] Zeh, D. W., & Smith, R. L. (1985). Paternal investment by terrestrial arthropods. American Zoologist, 25(3), 785-805.
[7] Gladstone, D. E. (1979). Promiscuity in monogamous colonial birds. The American Naturalist, 114(4), 545-557.
[8] Forbes, S. H., Adams, R. M., Bitney, C., & Kukuk, P. F. (2002). Extended parental care in communal social groups. Journal of Insect Science, 2(1), 22.
[9] Kullmann, E. J. (1972). Evolution of social behavior in spiders (Araneae ; Eresidae and Theridiidae). American Zoologist, 12(3), 419-426.
[10] Pas au sens évolutif, mais au niveau de la complexité des comportements et de la division du travail. Ce terme devrait être évité mais il est malheureusement très présent dans la littérature sur ce sujet.
[11] Wilson, E. O., & Holldobler, B. (2009). The superorganism : The beauty, elegance, and strangeness of insect societies. W. W Norton & Company, New York.
[12] Kim, K. W., Krafft, B., & Choe, J. C. (2005). Cooperative prey capture by young subsocial spiders. Behavioral Ecology and Sociobiology, 59(1), 92-100.
[13] Burch, T. L. (1979). The importance of communal experience to survival for spiderlings of Araneus diadematus (Araneae : Araneidae). Journal of arachnology, 1-18.
[14] Matheson, T., Rogers, S. M., & Krapp, H. G. (2004). Plasticity in the visual system is correlated with a change in lifestyle of solitarious and gregarious locusts. Journal of neurophysiology, 91(1), 1-12.
[15] Barron, A. B., Oldroyd, B. P., & Ratnieks, F. L. (2001). Worker reproduction in honey-bees (Apis) and the anarchic syndrome : a review. Behavioral Ecology and Sociobiology, 50(3), 199-208.
[16] Giron, D., Dunn, D. W., Hardy, I. C., & Strand, M. R. (2004). Aggression by polyembryonic wasp soldiers correlates with kinship but not resource competition. Nature, 430(7000), 676.
[17] Duffy, J. E. (1996). Eusociality in a coral-reef shrimp. Nature, 381(6582), 512.
[18] Fryxell, J. M. (1991). Forage quality and aggregation by large herbivores. The American Naturalist, 138(2), 478-498.
[19] Wilson, E. O. (2005). Kin selection as the key to altruism : its rise and fall. Social research, 159-166.
[20] Hamilton, W. D. (1964). The genetical evolution of social behaviour. I. Journal of theoretical biology, 7(1), 1-16.
[21] Smith, J. M. (1964). Group selection and kin selection. Nature, 201, 1145-1147.
[22] Nowak, M. A., Tarnita, C. E., & Wilson, E. O. (2010). The evolution of eusociality. Nature, 466(7310), 1057.
[23] Winston, M. L. (1991). The biology of the honey bee. harvard university press.
[24] Moroa, D., Witek, M., & Woyciechowski, M. (2008). Division of labour among workers with different life expectancy in the ant Myrmica scabrinodis. Animal Behaviour, 75(2), 345-350.